ABSTRACT
Sustainable forest management is an important strategy for the conservation of tropical secondary forests. However, knowledge about their potential for timber production is scarce, especially in the Central Amazon. We performed a silvicultural diagnosis in Central Amazon secondary forests to understand their potential for forest management. The diagnosis was carried out in two secondary forest patches at advanced successional stages located 38 km north and 75 km northeast of Manaus. We quantified the commercial tree species, classified their stem quality and growth conditions (crown exposure and liana infestation), and related tree size (diameter at breast height-DBH) to growth conditions. We identified 46 commercial tree species. Goupia glabra, a very important timber species, exhibited the highest density of individuals (more than 95 individuals ha-1). Good and reasonable quality stems accounted for 87% of the commercial trees, with 45 and 42% classified as of good and reasonable quality, respectively. The species composition and stem quality of the examined secondary forests support timber production through sustainable management. Commercial trees predominantly had crowns with low light exposure (63% with a Dawkins index lower than 4) and were liana infested (62% with lianas on some part of the tree), requiring silvicultural treatments to improve growth conditions. The number of trees under suppressed growth conditions (low light and liana-infested) varied according to species and tree size, requiring specific recommendations for silvicultural treatments. These results demonstrate the potential of secondary forests for timber production and provide valuable insights for management strategies aiming at promoting sustainable management to prevent secondary forest deforestation.
Keywords:
Regenerating forests; Alternative timber sources; Vegetation structure; Dominant species; Assisted natural regeneration.
RESUMO
O manejo florestal sustentável é uma estratégia importante para a conservação de florestas tropicais secundárias. No entanto, o conhecimento sobre o potencial dessas florestas para a produção de madeira é escasso, especialmente na Amazônia Central. Nós realizamos o diagnóstico silvicultural em florestas secundárias da Amazônia Central para analisar seu potencial para o manejo florestal. O diagnóstico foi conduzido em duas florestas secundárias em estágio avançado de regeneração, localizadas a 38 km ao norte e 75 km ao nordeste de Manaus. Nós quantificamos as espécies arbóreas comerciais, classificamos a qualidade do fuste e condições de crescimento (exposição da copa e infestação por lianas) de seus indivíduos e relacionamos o tamanho dos indivíduos com as condições de crescimento. Nós encontramos 46 espécies arbóreas comerciais. Goupia glabra, uma espécie madeireira muito importante, teve a maior densidade (acima de 95 indivíduos ha-1). 87% dos indivíduos das espécies comerciais possuem fustes de qualidade boa (45%) ou razoável (42%). As florestas secundárias estudadas possuem composição de espécies e qualidade de fustes que apoiam a produção de madeira através do manejo sustentável. Predominantemente, os indivíduos possuem copas pouco expostas à luz (63% delas com Índice de Dawkins abaixo de 4) e estavam infestados por lianas (62% com lianas em alguma parte da árvore), demandando tratamentos silviculturais para melhorar as condições de crescimento. A quantidade de indivíduos em condições suprimidas de crescimento (pouco iluminadas e infestadas com lianas) variou de acordo com as espécies e o tamanho do indivíduo, embasando recomendações específicas para tratamentos silviculturais. Esses resultados demonstram o potencial de florestas secundárias para a produção de madeira e fornecem valiosas informações para a formulação de estratégias de manejo, buscando promover o manejo sustentável como forma de evitar o desmatamento de florestas secundárias.
Palavras-chave:
Florestas em regeneração; Fontes alternativas de madeira; Estrutura da vegetação; Espécies dominantes; Regeneração natural assistida.
INTRODUCTION
Secondary forests are representative landscapes in the Brazilian Amazon, accounting for more than 16 million hectares and occupying approximately 20% of previously deforested areas (INPE 2021). Conserving these secondary forests is of the utmost urgency for Brazil due to their ecological importance in providing ecosystem services that mitigate the climate impacts of deforestation (Aliança pela Restauração na Amazônia 2024). The regeneration process enables the recovery of forest functionality and essential ecosystem services, such as carbon sequestration, biodiversity, hydrological and nutrient cycling, soil protection, and wildlife shelter (Chazdon 2014; Poorter et al. 2021; Heinrich et al. 2023). Additionally, conserving existing secondary forests and stimulating natural regeneration are low-cost strategies for forest restoration (REGENERA-Amazônia 2023). In this respect, encouraging secondary forest conservation may result in Brazil’s achieving the forest restoration goal of 12 million hectares, established in the Paris Agreement (Aliança pela Restauração na Amazônia 2024).
Despite their importance, secondary forests have the highest deforestation rates in the Brazilian Amazon (Nunes et al. 2020; Wang et al. 2020), with a more than 8% decline between 2014 and 2020 (INPE 2021). One of the reasons for deforestation is the lack of knowledge about the potential of secondary forests in generating income, prompting owners to replace them with other land uses (Pereira and Vieira 2001). In the Central Amazon, socioeconomic changes over time have intensified swidden agriculture, resulting in secondary forest degradation due to shorter fallow periods (Jakovac et al. 2016). One way to encourage owners to conserve their secondary forests is by demonstrating their economic potential through sustainable forest management (Schwartz et al. 2015; Bieng et al. 2021).
Sustainable secondary forest management requires silvicultural diagnosis to determine suitable silvicultural systems (Lamprecht 1990; Pancel 2016). This involves assessing the objectives and/or requirements of the population, site conditions, and secondary forest characteristics (Hutchinson 1991; Pancel 2016). Estimates indicate an increasing global demand for timber (FAO 2009; Barua et al. 2014), Brazil being one of the main exporters of tropical timber from old-growth forest management (Richardson and Peres 2016). However, under current forest management policies, timber stocks are bound to decline, thereby threatening sustainability (Piponiot et al. 2019; Sist et al. 2021). Sustainable secondary forest timber production is emerging as a means to foster their conservation and alleviate pressure on old-growth forests (Bieng et al. 2021; Sist et al. 2021).
Timber production from tropical secondary forest management is based on their capacity to quickly recover the density of commercial tree species (Finegan 1992; Doua-Bi et al. 2021; Zambiazi et al. 2021). However, studies showing the potential of Amazon secondary forests for timber production are scarce and concentrated in the Eastern Amazon (Alvino et al. 2005; Schwartz 2007), and nonexistent in the Central Amazon. Conducting a silvicultural diagnosis of Central Amazon secondary forest characteristics, including floristic composition, stem quality, and growth conditions of desirable species, is the initial step in assessing their suitability for timber provision and prescribing suitable silvicultural systems and treatments (Lamprecht 1990; Hutchinson 1991; Pancel 2016).
The need for silvicultural treatments may differ depending on the species and size of the individual. For example, in a late secondary forest, short- and long-lived pioneers dominate the canopy, while partially shade-tolerant species typically occupy subcanopy positions and thus have different available light levels (Finegan 1992; Chazdon 2014). With respect to tree size, as diameter increases, so does height (Feldpausch et al. 2011) and, consequently, the level of crown exposure to light (Poorter et al. 2005). The light level received by the crown is related to growth rates in secondary forests and may indicate priority individuals for silvicultural treatments (Mendoza-Hernández et al. 2019). Thus, analyzing how growth conditions are related to species and tree size helps establish priority species and individuals that will benefit from silvicultural treatments.
The central region is the second highest timber-producing region of the Amazonas state (Lentini et al. 2021), where secondary forests around 30 years old - denominated here as “late secondary forests” sensu Hartshorh (1980) are common (Silva-Junior et al. 2020). This is interesting because regeneration time is positively related to commercial tree species recovery (Doua-Bi et al. 2021), therefore these late secondary forests generate good expectations regarding the potential for timber supply. Therefore, assessing the potential of secondary forests for timber production in this region is an important strategy in devising alternative income sources for owners and encouraging their conservation. We performed a silvicultural diagnosis of two late secondary forests in the central region of the Amazonas state aimed at: 1) analyzing if secondary forests exhibit the species composition and structure needed for timber production; 2) evaluating the stem quality and growth conditions of commercial tree species in secondary forests; 3) analyzing whether tree size and growth conditions are associated.
MATERIAL AND METHODS
Study site
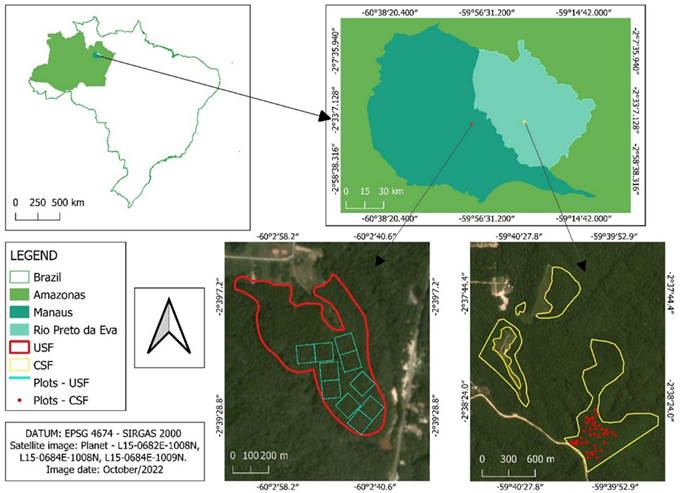
This study was conducted in two tropical secondary forest patches in the Central Amazon, one on the Experimental Farm of the Federal University of Amazonas - FAEXP/UFAM (02°38’S, 60°03.5’W) 38 km north of Manaus, and the other on Carapanã-Açu Farm (2°38’S; 50°40’W), 75 km northeast of Manaus in Rio Preto da Eva, both in the Amazonas state, Brazil. The straight-line distance between the two patches is approximately 42 km (Supplementary Material, Figure S1). The climate in the region is Af, according to the Köppen-Geiger classification system (Alvares et al. 2013), and the surrounding vegetation is predominantly tropical moist forest (IBGE 2012). According to a historical data series of weather stations in Manaus (1988 - 2018), average annual rainfall in the region is 2366 mm, monthly average temperature ranges from 26.6 to 28.7°C, and average relative humidity from 74 to 86% (INMET 2023). The two secondary forest patches are established on plateaus, consisting of well-drained nutrient-poor clay oxisols with high aluminum concentrations (Chauvel 1982).
Secondary forests characterization
The secondary forest at FAEXP/UFAM, hereinafter denominated USF, occupies an area of 35 ha. Between 1984 and 1986, the old-growth forest was cut down and burned to implement monocultures, such as Brazil nut (Bertholletia excelsa Bonpl.) and peach palms (Bactris gasipaes Kunth), but the area was abandoned immediately after planting, making secondary vegetation regrowth possible (Supplementary Material, Figure S2). We observed some living B. excelsa trees, but avoided these areas and focused on secondary forests with a homogeneous stand age of 35 years (with reference to 2021). Prior to cutting and burning, the area was selectively logged, allowing light-demanding species to regenerate in the gaps. If the young individuals were not efficiently cut or survived the fire, there may be individuals older than the estimated age of the forest. The secondary forest at the Carapanã-Açu Farm, hereinafter denominated CSF, occupies an area of 50 ha. In 1985, the vegetation was clear-cut and burned, and the soil was plowed and harrowed for rubber tree planting (Hevea brasiliensis Mull. Arg.), but the area was abandoned immediately after planting (Supplementary Material, Figure S2), and the seedlings died due to a lack of stand tending. Thus, at the time of the study (2021), the secondary forest was 36 years old.
Data collection
In 2021, we installed five experimental blocks with two 1-ha plots at UFS, totaling ten hectares of inventories. We installed these blocks for an experiment of secondary forest improvement through liberation thinning, and arranged them to ensure that the plots were at the same elevation (elevation range = 75 to 113 m) (Supplementary Material, Figure S1). We carried out taxonomical identification and measured the diameter at breast height (DBH) of all individuals with DBH ≥ 10 cm in the plots. We listed all commercial species with potential for timber production, based on previously developed lists of commercial timber species from the Amazon (Cysneiros et al. 2018; IBAMA 2019; Andrade et al. 2022a; ITTO 2022; LPF 2022). We also included species that are not recognized by the timber market but whose timber are used by the local communities and regenerate at high density in the forest. The species included are Vismia cayennensis, Croton matourensis and Sacoglottis guianensis. Details about the potential use of these species are in the Supplementary Material (Table S1). All individuals of the commercial species were classified for stem quality and growth conditions. Individuals were visually classified by the same person to avoid subjectivity. Stem quality indicate the degree of tortuosity and bifurcation, and growth conditions indicate the degree of suppression by surrounding trees (assessed by canopy exposure) and lianas (assessed by liana infestation). The classes for stem quality, crown exposure, and liana infestation used are detailed and referenced in Table 1.
At CSF, also in 2021, we performed a silvicultural analysis of the Goupia glabra and Vismia cayennesis because previous analyses showed that these species were the most abundant of the commercial species at USF. Therefore, we conducted this sampling to determine if the patterns found for these species at USF were replicated at another secondary forest in the region. We installed 42 10 x 10 m plots, totaling 0.42 hectares of inventories (Supplementary Material, Figure S1), and measured the DBH of all individuals with DBH ≥ 10 cm in the plots. Only Goupia glabra and Vismia cayennensis individuals were botanically identified and classified according to stem quality and growth conditions (Table 1).
Data analysis
To determine whether the secondary forests exhibit the species composition and structure needed for timber production, we conducted the following analysis: At USF, we first quantified the number of commercial tree species, and then calculated the mean and 95% confidence interval of the total basal area and density of the forest and commercial tree species. For each commercial tree species, we calculated the density (De), dominance (Do) and frequency (Fr) in absolute (ab) and relative (re) values. Next, we calculated their importance [IV = (De relative + Do relative + Fr relative) / 3] and coverage values [CV = (De relative + Do relative) / 2]. We then evaluated the diameter distribution of the five dominant commercial tree species and other commercial tree species grouped (hereinafter denominated OCTS) through histograms. OCTS individuals were grouped for analyses because these species had low densities (less than 10 individuals ha-1) and were absent from some plots, resulting in insufficient samples for some analyses. These species also had similar characteristics, with most individuals exhibiting small diameters and low crown exposure. The five dominant species and OCTS are shown in the Results section (Table 2). At CSF, we calculated the mean and 95% confidence interval of the total basal area and density, and the De (ab and re), Do (ab and re), Fr (ab), CV, and diametric distribution of G. glabra and V. cayennensis.
Horizontal structure of commercial species regenerating in FAEXP/UFAM (USF) and Carapanã-Açu Farm (CSF) secondary forests, with species ranked from highest to lowest importance value. Do - Dominance, De - Density, Fr - Frequency, IV - Importance Value, CV - Coverage value, ab - absolute, re - relative.
We assessed the stem quality and growth conditions of the commercial tree species in the secondary forests as follows: At USF, we first quantified the density of individuals in each stem quality and growth conditions (crown exposure and liana infestation) class, in each plot, for the five dominant commercial tree species and the OCTS. We then reported the mean and standard deviation of the density of individuals in each class of stem quality and growth conditions. At CSF, we calculated the density of G. glabra and V. cayennensis individuals in each stem quality and growth conditions classes.
We analyzed if tree size and growth conditions are associated through GLMMs. The models were applied at the individual level, with DBH as the dependent variable and crown exposure or liana infestation as independent variables. Secondary forests (USF and CSF) and species were used as random factors, as well as gamma distribution and the logarithmic link function. All statistical analyses were performed with R statistical software, version 4.2.1. (R Core Team 2022) and the GLMMs applied using the glmmTMB package.
RESULTS
Floristic composition and horizontal structure of the commercial tree pool
At USF, the total basal area (for trees ≥ 10 cm DBH) was 16.94 ± 0.71 m2 ha-1 and the density 582 ± 19 individuals ha-1. We found 254 species regenerating naturally, 46 of which are commercial tree species. The total basal area of the commercial tree species (for trees ≥ 10 cm DBH) was 8.63 ± 1.07 m2 ha-1 and the density 281 ± 34 individuals ha-1. Commercial tree species accounted for 41 and 49% of the importance and coverage values, respectively. Goupia glabra, Vismia cayennensis, Guatteria punctata, Croton matourensis and Laetia procera were the five dominant commercial tree species. These species occur in high densities (more than 18 individuals ha-1), but G. glabra and V. cayennensis stand out for their very high densities (110 and 61 individuals ha-1, respectively) (Table 2). The diameter distribution patterns varied among species, with a reverse J-shaped diameter distribution curve for G. glabra, G. punctata and the OCTS, while V. cayennensis, C. matourensis, and L. procera exhibited similar bell-shaped curves. Only OCTS had individuals with DBH greater than or equal to 50 cm, with fewer than one percent of individuals in these classes (Figure 1).

Percentage of individuals in each diameter class for five dominant commercial species and other commercial tree species - OCTS (others) at USF (black), and G. glabra and V. cayennensis at CSF (gray).
At CSF, the total basal area (for trees ≥ 10 cm DBH) was 18.61 ± 1.88 m2 ha-1 and the density 590 ± 42 individuals ha-1. G. glabra and V. cayennensis accounted for 34% of the coverage value and had densities of 95 and 62 individuals ha-1, respectively (Table 2). G. glabra had a similar reverse J-shaped diameter distribution curve, and V. cayennensis a bell-shaped curve (Figure 1).
Stem quality, crown exposure and liana infestation of the commercial species
When the commercial tree species from both secondary forests were summed, we found that 45% exhibited good (class 3), 42% reasonable (class 2) and 13% poor (class 1) stem quality. At USF, classes 3, 2, and 1 represent 45, 42, and 13% of individuals of commercial species, respectively, while at CSF they represent 49, 21, and 30%. Overall, commercial tree species had low crown exposure and were infested by lianas, although the crown exposure pattern varied among species. Considering all the commercial tree species from both sites, 63% had crowns with low light exposure (Dawkins index lower than 4), and 62% contained lianas on some part of the tree (were in classes 2, 3 or 4 of liana infestation).
At USF, G. glabra and V. cayennensis had the highest number of individuals with reasonable stem quality, while G. punctata, C. matourensis, L. procera, and OCTS had the highest number with good stem quality (Figure 2a). With respect to crown exposure, G. glabra and OCTS had more individuals in class 2 and 3, while V. cayennensis, G. punctata, C. matourensis, and L. procera had more individuals in class 4 (fully exposed vertically) (Figure 2b). The percentage of G. glabra individuals with lianas on some parts of the tree was 75%, for V. cayennensis 58%, G. punctata 63%, C. matourensis 58%, L. procera 50% and OCTS 60% (Figure 2c).

Density of individuals in each stem quality (a), crown exposure (b), and liana infestation (c) class for five dominant commercial species and other commercial tree species - OCTS at USF. Bars represent the means, and error bars the standard deviation. For stem quality, crown exposure and liana infestation classes, see Table 1.
At CSF, 50% of G. glabra individuals had good, 19% reasonable, and 31% poor stems. For V. cayennensis, 48% of individuals had good, 24% reasonable and 28% poor stems. Most G. glabra individuals had crowns fully exposed to light vertically, but a high percentage exhibited crowns partially exposed vertically. Most V. cayennensis individuals had crowns partially exposed vertically, but a high percentage displayed crowns fully exposed vertically and fully exposed vertically and laterally, while 65 and 71% of G. glabra and V. cayennensis individuals, respectively, had lianas on some parts of the tree (Figure 3).

Total density of individuals in each stem quality, crown exposure, and liana infestation class for G. glabra and V. cayennensis at CSF. For stem quality, crown exposure and liana infestation classes, see Table 1.
Tree size and growth conditions associations
There was a significant association between crown exposure and liana infestation and DBH (Figure 4). Crown exposure and DBH were positively associated, that is, larger trees had progressively higher crown exposure. The mean DBH of the highest crown exposure (class 5) was 2.2-fold higher than that of the lowest (class 1). Mean DBH values changed between liana infestation levels but to a small degree. The highest mean DBH was observed in class 4, being 1.2-fold higher than the level with the lowest (class 2).

Diameter at breast height (DBH) of commercial tree species on different conditions of crown exposure (a) and liana infestation (b). Bars represent the means of DBH, and error bars the standard deviation. GLMM p values are expressed as * < 0.05; ** < 0.01; *** < 0.001. Number of samples = 2609. For the crown exposure and liana infestation classes, see Table 1
DISCUSSION
Late secondary forests are rich and dense in commercial timber species
Commercial tree species are important components of forest structure, as reflected by their high density of individuals, dominance, frequency, importance and coverage values. Commercial tree species density at USF exceeded 152 individuals ha-1 (for DBH ≥ 10), which is the amount typically exploited for timber in old-growth forests of the Southwestern Amazon (Selaya et al. 2017). The density of commercial tree species at USF also exceeded the 105 commercial timber species ha-1 (for DBH ≥ 15 cm) found in an old-growth forest in the Central Amazon (Souza 2015). However, this cannot be expected for all secondary forests in the region. The recovery of species richness after disturbance is dependent on distance from propagule sources, land use history, the presence of remnant trees, and regeneration time (Mesquita et al. 2001; Doua-Bi et al. 2021; Jakovac et al. 2021). USF and CSF are close to the remaining old-growth forest (Supplementary Material, Figure S1), were subjected to previous low intensity land use with only one cut-burn cycle, and have been regenerating for a long time (Supplementary Material, Figure S2). These conditions contribute to commercial tree species recovery (Doua-Bi et al. 2021).
The dominant species in our studied sites (G. glabra) is one of the most commercialized native timber species in Brazil (IBAMA 2019; Andrade et al. 2022a), demonstrating the potential of secondary forests for timber production. The next dominant commercial tree species in the rank (V. cayennensis, G. punctata, C. matourensis, and L. procera) are less recognized by the timber market, but their inclusion meets the current need to expand the range of species in forest management (Sist et al. 2021; Andrade et al. 2022b). Scientific knowledge of wood technology to propose uses for these species is still required to achieve market acceptance. However, L. procera is relatively well studied and has recently been proposed to meet the demand for Amazonian timber (Andrade et al. 2022b).
The wood density of commercial tree species in secondary forests varies, resulting in different possible applications. G. glabra, C. matourensis, and L. procera have moderately heavy timber, with wood densities of 0.73, 0.62, and 0.65 g cm-3, respectively. V. cayennensis and G. punctata have light timber, with respective wood densities of 0.49 and 0.46 g cm-3 (Zanne et al. 2009). G. glabra is recommended for heavy external and internal construction, tool handles, truck bodies, and shipbuilding (Gurgel et al. 2015; ITTO 2022); C. matourensis for linings, toys, and boxes (Loureiro 1968); L. procera for external and internal construction, furniture, and floors (Finegan 1992; Andrade et al. 2022b); V. cayennensis for internal construction (information provided by the local community) and G. punctata for internal construction, household items, toys, and boxes (LPF 2022).
Stem quality and growth conditions
The stems of commercial tree species were mostly good or reasonable, a critical factor for achieving good usable stem volume free of branches, heavy tortuosity, and bifurcation (Köhl and Marchetti 2016). This attribute is fundamental in achieving a profitable roundwood volume in subsequent logging operations. Most commercial tree species were under low light and liana-infested, indicating suppressed growth. Trees with limited crown exposure intercept less light, resulting in reduced photosynthetic rates and consequently slow growth (Peña-Claros et al. 2008; Kenzo et al. 2015; Mendoza-Hernández et al. 2019). Lianas compete with trees for light, water, and nutrients, resulting in reduced growth and, in some cases, death (Reis et al. 2020). From a timber production perspective, the lower growth caused by competition of canopy trees and lianas delays logging, requiring silvicultural treatments to improve growth conditions and management profitability (Penã-Claros et al. 2008; Putz et al. 2023).
The crowns of V. cayennensis, G. punctata, C. matourensis, and L. procera were more exposed to light than those of G. glabra and OCTS individuals, which may be related to the ecological succession during regeneration. At the onset of regeneration, short-lived pioneers dominate the canopy, and are then gradually replaced by long-lived pioneers and partially shade-tolerant individuals (Chazdon 2014). V. cayennensis, C. matourensis, and L. procera had a diameter distribution typical of pioneer species that regenerate in the first stage of succession, grow rapidly, and soon leave the small diameter classes. In addition, the juvenile mortality of pioneers caused by shading also contributes to the low density of individuals in the small diameter classes and population decline (Turner 2001). Field observations revealed the natural mortality of V. cayennensis and C. matourensis (Supplementary Material, Figure S3). Although L. procera exhibited typical pioneer diameter distribution, it also contained many individuals in the lowest diameter class. Efficient height growth rates may lead this species to compete effectively for light and persist longer in the secondary forest (Turner 2001). The same efficient height growth rates may explain the occurrence of most G. punctata individuals with well-lit crowns (Supplementary Material, Figure S4), even with the typical diameter distribution of late successional species (J-shaped).
Low diameter trees were typically small and located below the canopy, reflecting suppressed growth that requires more silvicultural intervention (Poorter et al. 2005; Feldpausch et al. 2011; Finegan 2015; Mendoza-Hernández et al. 2019). The largest trees had slightly more lianas, because they are older and have larger crowns, meaning lianas have more time to accumulate and more crown area to occupy (Alvira et al. 2004). Alternatively, the high crown exposure of the largest trees provides lianas with the light they need (Venegas-González et al. 2020). Thus, even if the larger trees are well lit, liana infestation can reduce timber yield.
Implications and recommendations for secondary forest management
The floristic composition and stem quality of the two evaluated Central Amazon secondary forests indicate good potential for timber production. The frequent occurrence of the commercial tree species found in other Amazonian secondary forests, especially G. glabra, is a valid reason for proposing secondary forests for timber management (Finegan 1996; Bentos et al. 2013; Bentos et al. 2014). The high density of some commercial tree species in secondary forests, greater than that found in old-growth Amazonian forests (Souza et al. 2015; Selaya et al. 2017), facilitates management in terms of product supply, making these forests a potentially valuable source of income for local communities. In secondary forests, the type and size of commercial tree species differ from those in old-growth forests. Secondary forests typically provide lighter and less valuable timber from pioneer species (Finegan 1992; Toledo-Aceves et al. 2022). In the present study, some of the commercial tree species have moderately heavy timber, and the dominant (G. glabra) has high value timber, representing a possible difference in terms of profitability for Central Amazon secondary forests.
The growth conditions of commercial tree species should be improved with silvicultural treatments in order to increase growth rates and anticipate logging cycles. Liberation thinning and liana cutting are recommended for desirable trees to reverse suppressed growth (Lamprecht 1990; Hu et al. 2020). Liberation thinning may be concentrated in small or medium diameter individuals and species that experience the worst growth conditions, such as G. glabra and OCTS in our study. On the other hand, liana cutting should be done whenever there is infestation, especially in larger trees. The use of timber from thinning can provide initial income for owners and partially support the application of silvicultural treatments. Thinning of declining high canopy layer species, such as V. caynennesis and C. matourensis, enhances light availability and provide a valuable timber. In addition, the light gaps created may favor their regeneration (Turner 2001), maintaining them in the forest for further logging cycles.
Further silvicultural diagnosis should be carried out at other sites to determine whether the high density of commercial tree species reported here is a general feature of central Amazonian secondary forests, and the factors that may be involved, such as land-use history (Doua-Bi et al. 2021; Jakovac et al. 2021). Secondary forest management should be facilitated by flexible legislation and incentive policies, and regulated to prevent ecosystem services from being threatened (Toledo-Aceves et al. 2021, 2022). Multiple secondary forest use should be considered as a complementary strategy to improve profitability. Payment for ecosystem services, management of non-timber products, and enrichment of gaps created by silvicultural treatments can provide quick economic returns, making secondary forest management more attractive to owners (Schwartz et al. 2015; Bieng et al. 2021; Toledo-Aceves et al. 2021).
CONCLUSIONS
The Central Amazon secondary forests studied here exhibit the species composition and density needed for sustainable forest management. Moreover, the stems of commercial species display suitable quality for roundwood production. However, a large proportion of these stems are found under limiting growth conditions, and silvicultural treatments are necessary to enhance growth rates and anticipate logging. Future studies should expand sampling efforts to other secondary forests, in addition to assessing the impact of silvicultural treatments on growth. These efforts are crucial in estimating logging cycles and establishing effective silvicultural systems.
ACKNOWLEDGMENTS
We acknowledge the Laboratório de Silvicultura of the Federal University of Amazonas, the Experimental Farm of the Federal University of Amazonas, the Laboratório de Manejo of the University of State of Amazonas, and the Carapanã-Açu Farm for support on logistics and fieldwork. We are thankful to the following sources of financial support: Edital PROCAD/CAPES AMAZÔNIA 2018 - Linha 2 (grant 88881.199859/2018-01); Chamada Pública 001/2020 - FAPESP/FAPEAM (grant 01.02.016301.00280/2021); Programa Institucional de Apoio à Pós-Graduação Stricto Sensu - POSGRAD/FAPEAM (Resolução: N. 006/2020, N. 008/2021, and 005/2022); Edital N. 001/2023 - UNIVERSAL - FAPEAM 20 ANOS (grant 01.02.016301.04320/2023-77). This study was partially funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001, and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). G. S. Modolo and D. Coelho Silva thank the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) for their scholarships. V. A. H. F. dos Santos thanks Universidade do Estado do Amazonas for the academic productivity grant (GPA UEA). M. J. Ferreira acknowledges the research productivity grant provided by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), grant 315005/2023-8.
REFERENCES
-
Aliança pela Restauração na Amazônia. 2024. POSICIONAMENTO - É urgente proteger a vegetação secundária na Amazônia. ( (https://aliancaamazonia.org.br/wp-content/uploads/2024/02/Alianca_posicionamento_veg_secundaria.docx.pdf ). Acessed on 20 may 2024.
» https://aliancaamazonia.org.br/wp-content/uploads/2024/02/Alianca_posicionamento_veg_secundaria.docx.pdf - Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. 2013. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift 22: 711-728.
- Alvino, F.O.; Silva, M.F.F.; Rayol, B.P. 2005. Potencial de uso das espécies arbóreas de uma floresta secundária, na Zona Bragantina, Pará, Brasil. Acta Amazonica 35: 413-420.
- Alvira, D.; Putz, F.E.; Fredericksen, T.S. 2004. Liana loads and post-logging liana densities after liana cutting in a lowland forest in Bolivia. Forest Ecology and Management 190: 73-86.
- Andrade, M.B.T.; dos Santos, H.; Nunes, F.; Costa, J.N.; Lentini, M.W. 2022a. Produção de madeira e diversidade de espécies arbóreas exploradas na amazônia brasileira: situação atual e recomendações para o setor florestal. Boletim Técnico Timberflow 8. Imaflora, Piracicaba. 19p.
- Andrade, M.B.T.; Zenid, G.J.; Lentini, M.W.; dos Santos, H.; Nunes, F.; Costa, J.N. 2022b. Espécies madeireiras nativas menos comercializadas: uma oportunidade para produção e conservação na Amazônia Brasileira. Boletim Técnico Timberflow 9. Imaflora, Piracicaba . 19p.
- Barua, S.K.; Lehtonen, P.; Pahkasalo, T. 2014. Plantation vision: potentials, challenges and policy options for global industrial forest plantation development. International Forestry Review 16: 117-127.
- Bieng, M.A.N.; Oliveira, M.S.; Roda, J.M.; Boissière, M.; Hérault, B.; Guizol, P.; et al. 2021. Relevance of secondary tropical forest for landscape restoration. Forest Ecology and Management 493: 119265.
- Bentos, T.V.; Nascimento, H.E.M.; Williamson, G.B. 2013. Tree seedling recruitment in Amazon secondary forest: Importance of topography and gap micro-site conditions. Forest Ecology and Management 287: 140-146.
- Bentos, T.V.; Mesquita, R.C.G.; Camargo, J.L.C.; Williamson, G.B. 2014. Seed and fruit tradeoffs - the economics of seed packaging in Amazon pioneers. Plant Ecology & Diversity 7: 371-382.
- Chauvel, A. 1982. Os latossolos amarelos, álicos, argilosos, dentro dos ecossistemas experimentais do INPA e da região vizinha. Acta Amazonica 12: 47-60.
- Chazdon, R.L. 2014. Second growth: The promise of tropical forest regeneration in an age of deforestation The University of Chicago Press, Chicago. 449p.
- Cysneiros, V.C.; Júnior, J.O.M.; Lanza, T.R.; de Moraes, J.C.R.; Samor, O.J.M. 2018. Espécies madeireiras da Amazônia: riqueza, nomes populares e suas peculiaridades. Pesquisa Florestal Brasileira 38: 1-14.
- Dawkins, H.C.; Field, D.R.B. 1978. A long-term surveillance system for British woodland vegetation Oxford University, Oxford. 106p.
- Doua-Bi, G.Y.; Zo-Bi, I.C.; Amani, B.H.; Elogne, A.G.; N’dja, J.K.; N’Guessan, A.E.; et al. 2021. Taking advantage of natural regeneration potential in secondary forests to recover commercial tree resources in Côte d’Ivoire. Forest Ecology and Management 493: 119240.
- FAO. 2009. State of the World’s Forests 2009. In FAO, Rome, Italy.
- Feldpausch, T.R.; Banin, L.; Phillips, O.L.; Baker, T.R.; Lewis, S.L.; Quesada, C.A.; et al. 2011. Height-diameter allometry of tropical forest trees. Biogeosciences 8: 1081-1106.
- Finegan, B. 1992. The management potential of neotropical secondary lowland rain forest. Forest Ecology and Management 47: 295-321.
- Finegan, B. 1996. Pattern and process in neotropical secondary rain forests: the first 100 years of succession. Trends in Ecology & Evolution 11: 119-124.
- Finegan, B. 2015. 21st century viewpoint on tropical silviculture. In: Pancel, L.; Kohl, M. (Eds.). Tropical Forestry Handbook Springer, Berlin Heidelberg. p. 605-1638.
- Gurgel, E.S.; Gomes, J.I.; Groppo, M.; Martins-da-Silva, R.C.V.; de Souza, A.S.; Margalho, L.; et al. 2015. Conhecendo Espécie de Plantas da Amazônia: Cupiúba (Goupia glabra Aubl. - Goupiaceae). Comunicado Técnico. EMBRAPA, Belém. 7p.
- Hartshorn, G.S. 1980. Neotropical forest dynamics. Biotropica 12: 23-30.
- Heinrich, V.H.A.; Vancutsem, C.; Dalagnol, R.; Rosan, T.M.; Fawcett, D.; Silva-Junior, C.H.L.; et al. 2023. The carbon sink of secondary and degraded humid tropical forests. Nature 615: 436-442.
- Hu, J.; Herbohn, J.; Chazdon, R.L.; Baynes, J.; Vanclay, J. 2020. Long-term growth responses of three Flindersia species to different thinning intensities after selective logging of a tropical rainforest. Forest Ecology and Management 476: 118442.
- Hutchinson, I.D. 1991. Diagnostic sampling to orient silviculture and management in natural tropical forest. Commonwealth Forestry Review 70: 113-132.
-
IBAMA. 2019. Instituto Brasileiro do Meio Ambiente e dos Recursos Renováveis. Produção madeireira de espécies nativas brasileiras: 2012 a 2017. 378p. https://drive.google.com/file/d/12wqidQwzN_nTEEoxaHAk-kcHqzy4zUsK/view
» https://drive.google.com/file/d/12wqidQwzN_nTEEoxaHAk-kcHqzy4zUsK/view - IBGE, 2012. Instituto Brasileiro de Geografia e Estatística - Manual técnico da vegetação brasileira (2 ed.). Rio de Janeiro. 271 p.
-
INMET. 2023. Instituto Nacional de Meteorologia - BDMEP - Banco de Dados Meteorológicos para Ensino e Pesquisa. ( (https://portal.inmet.gov.br/dadoshistoricos ). Accessed on 6 Nov 2023.
» https://portal.inmet.gov.br/dadoshistoricos -
INPE. 2021. Instituto Nacional de Pesquisas Espaciais - Monitoramento do uso e cobertura da terra nas áreas desflorestadas da Amazônia Legal - TerraClass Amazônia. Nota Técnica - Estimativa da área de Vegetação Secundária na Amazônia Legal (2020). ( (https://www.gov.br/inpe/pt-br/assuntos/ultimas-noticias/nota-tecnica-2013-estimativa-da-area-de-vegetacao-secundaria-na-amazonia-legal-2020 ). Accessed on 1 Feb 2023.
» https://www.gov.br/inpe/pt-br/assuntos/ultimas-noticias/nota-tecnica-2013-estimativa-da-area-de-vegetacao-secundaria-na-amazonia-legal-2020 -
ITTO. 2022. International Tropical Timber Organization - Lesser Used Species. ( (http://www.tropicaltimber.info/pt-br/ ). Accessed on 1 Feb 2023.
» http://www.tropicaltimber.info/pt-br/ - Jakovac, C.C.; Peña-Claros, M.; Mesquita, R.C.G.; Bongers, F.; Kuyper, T.W. 2016. Swiddens under transition: Consequences of agricultural intensification in the Amazon. Agriculture, Ecosystems & Environment 218: 116-125.
- Jakovac, C.C.; Junqueira, A.B.; Crouzeilles, R.; Peña-Claros, M.; Mesquita, R.C.G.; Bongers, F. 2021. The role of land-use history in driving successional pathways and its implications for the restoration of tropical forests. Biological Reviews 96: 1114-1134.
- Kenzo, T.; Inoue, Y.; Yoshimura, M.; Yamashita, M.; Tanaka-Oda, A.; Ichie, T. 2015. Height-related changes in leaf photosynthetic traits in diverse Bornean tropical rain forest trees. Oecologia 177: 191-202.
- Köhl, M., Marchetti, M. 2016. Measurements and Assessments on Field Plots. In: Pancel, L.; Kohl, M. (Eds.). Tropical Forestry Handbook Springer, Berlin Heidelberg . p. 729-731.
-
LPF. 2022. Laboratório de Produtos Florestais - Banco de dados de madeiras brasileiras. Serviço Florestal Brasileiro. ( (https://lpf.florestal.gov.br/pt-br/madeiras-brasileiras ). Accessed on 1 Feb 2023.
» https://lpf.florestal.gov.br/pt-br/madeiras-brasileiras - Lamprecht, H. 1990. Silvicultura nos trópicos: Ecossistemas florestais e respectivas espécies arbóreas - possibilidades e métodos de aproveitamento sustentado Instituto de Silvicultura da Universidade Gottingen, GTZ, Eschborn. 343p.
- Lentini, M.; Andrade, M.; Nunes, F.; dos Santos, H. 2021. Amazonas, o gigante florestal brasileiro: Desafios e oportunidades para o uso e a conservação das florestas naturais de produção. Boletim Técnico Timberflow 5. Imaflora, Piracicaba . 20p.
- Loureiro, A.A. 1968. Contribuição ao estudo anatômico de Croton lanjouwensis (Muell Arg.) Jablonski e Croton matourensis Aublet (Euphorbiaceae). Manaus, CNP/INPA nº 24. 16p.
- Mendoza-Hernández, M.; Gerez-Fernández, P.; Purata-Velarde, S.; Todelo-Aceves, T. 2019. Growth rates of valuable tree species in secondary tropical montane cloud forests in Mexico: influence of tree size, crown position and competition. Madera y Bosques 25: 1-11.
- Mesquita, R.C.G.; Ickes, K.; Ganade, G.; Williamson, G.B. 2001. Alternative successional pathways in the Amazon Basin. Journal of Ecology 89: 528-537.
- Nunes, S.; Oliveira Jr, L.; Siqueira, J.; Morton, D.C.; Souza Jr, C.M. 2020. Unmasking secondary vegetation dynamics in the Brazilian Amazon. Environmental Research Letters 15: 034057.
- Pancel, L. 2016. Technical Orientation of Silviculture in the Tropics. In: Pancel, L.; Kohl, M. (Eds.). Tropical Forestry Handbook Springer, Berlin Heidelberg . p. 1640-1687.
- Peña-Claros, M.; Fredericksen, T.S.; Alarcón, A.; Blate, G.M.; Choque, U.; Leaño, C.; et al. 2008. Beyond reduced-impact logging: Silvicultural treatments to increase growth rates of tropical trees. Forest Ecology and Management 256: 1458-1467.
- Pereira, C.A.; Vieira, I.C.G. 2001. A importância das florestas secundárias e os impactos de sua substituição por plantios mecanizados de grãos na Amazônia. Interciencia 26: 337-341.
- Piponiot, C.; Rödig, E.; Putz, F.E.; Rutishauser, E.; Sist, P.; Ascarrunz, N.; et al. 2019. Can timber provision from Amazonian production forests be sustainable? Environmental Research Letters 14: 064014.
- Poorter, L.; Bongers, F.; Sterck, F.J.; Wöll, H. 2005. Beyond the regeneration phase: differentiation of height-light trajectories among tropical tree species. Journal of Ecology 93: 256-267.
- Poorter, L.; Craven, D.; Jakovac, C.C.; van der Sande, M. T.; Amissah, L.; Bongers, F.; et al. 2021. Multidimensional tropical forest recovery. Science 374: 1370-1376.
- Putz, F.E.; Cayetano, D.T.; Belair, E.P.; Ellis, P.W.; Roopsind, A.; Griscom, B.W.; et al. 2023. Liana cutting in selectively logged forests increases both carbon sequestration and timber yields. Forest Ecology and Management 539: 121038.
- R Core Team. 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing.
- Richardson, V.A.; Peres, C.A. 2016. Temporal decay in timber species composition and value in Amazonian logging concessions. PLoS ONE 11: e0159035.
- REGENERA-Amazônia. 2023. Recomendações para o monitoramento da regeneração natural na Amazônia. Manaus, 24p.
- Reis, S.M.; Marimon, B.S.; Morandi, P.S.; Elias, F.; Esquivel-Muelbert, A.; Junior, B.H.M.; et al. 2020. Causes and consequences of liana infestation in southern Amazonia. Journal of Ecology 108: 2184-2197.
- Sant’Ana, V.Z.; Freitas, M.L.M.; de Moraes, M.L.T.; Zanata, M.; Zanatto, A.C.S.; de Moraes, M.A.; et al. 2013. Parâmetros genéticos em progênies de polinização aberta de Enterolobium contortisiliquum (Vell.) Morong em Luiz Antonio, SP, Brasil. Hoehnea 40: 515-520.
- Schwartz, G. 2007. Manejo sustentável de florestas secundárias: espécies potenciais no nordeste do Pará, Brasil. Amazônia: Ciência & Desenvolvimento 3: 125-147.
- Schwartz, G.; Ferreira, M.S.; Lopes, J.C. 2015. Silvicultural intensification and agroforestry systems in secondary tropical forests: a review. Amazonian Journal of Agricultural and Environmental Sciences 58: 319-326.
- Selaya, N.G.; Zuidema, P.A.; Baraloto, C.; Vos, V.A.; Brienen, R.J.W.; Pitman, N.; et al. 2017. Economically important species dominate aboveground carbon storage in forests of southwestern Amazonia. Ecology and Society 22: 40.
- Silva-Junior, C.H.L.; Heinrich, V.H.A.; Freire, A.T.G.; Broggio, I.S.; Rosan, T.M.; Doblas, J.; et al. 2020. Benchmark maps of 33 years of secondary forest age for Brazil. Scientific Data 7: 269.
- Sist, P.; Piponiot, C.; Kanashiro, M.; Peña-Claros, M.; Putz, F.E.; Schulze, M.; et al 2021. Sustainability of Brazilian forest concessions. Forest Ecology and Management 496: 119440.
-
Souza, M.A.S. 2015. Dinâmica e produção de uma floresta sob regime de manejo sustentável na Amazônia Central. Master´s dissertation. Universidade Federal do Amazonas (UFAM), Brazil, 102p. (https://tede.ufam.edu.br/handle/tede/4680)
» https://tede.ufam.edu.br/handle/tede/4680 - Toledo-Aceves, T.; Guariguata, M.R.; Günter, S.; Porter-Bolland, L.; Merino, L. 2021. Overcoming key barriers for secondary cloud forest management in Mexico. Land 10: 1078.
- Toledo-Aceves, T.; Günter, S.; Guariguata, M.R.; García-Díaz, M.; Zhunusova, E. 2022. Financial revenues from timber harvesting in secondary cloud forests: A case study from Mexico. Forests 13: 1496.
- Turner, I.M. 2001. The Ecology of Trees in the Tropical Rain Forest Cambridge University Press, Cambridge. 298p.
- Wang, Y.; Ziv, G.; Adami, M.; de Almeida, C.A.; Antunes, J.F.G.; Coutinho, A.C.; et al. 2020. Upturn in secondary forest clearing buffers primary forest loss in the Brazilian Amazon. Nature Sustainability 3: 290-295.
- Venegas-González, A.; Mello, F.N.A.; Schnitzer, S.A.; César, R.G.; Tomazello-Filho, M. 2020. The negative effect of lianas on tree growth varies with tree species and season. Biotropica 52: 836-844.
- Zambiazi, D.C.; Fantini, A.C.; Piotto, D.; Siminski, A.; Vibrans, A.C.; Oller, D.C.; et al. 2021. Timber stock recovery in a chronosequence of secondary forests in Southern Brazil: Adding value to restored landscapes. Forest Ecology and Management 495: 119352.
-
Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; et al. 2009. Global wood density database. Dryad. (http://hdl.handle.net/10255/dryad.235).
» http://hdl.handle.net/10255/dryad.235
-
CITE AS:
Modolo, G.S; Santiago, D.A; Dos Santos, V.A.H.F; Guimarães, Z.T.M; Campos, T.F.L; Reis, A.F; Coelho Silva, D; Tomazello Filho, M; Schöngart, J; Albiero Junior, A; Assis Pereira, G; Ferreira, M.J. 2025. Assessing the potential of tropical secondary forests in the Central Amazon for timber production. Acta Amazonica 55: e55ag24002.
Data availability
The data that support the findings of this study are available, upon request, from the corresponding author, Marciel José Ferreira.
SUPPLEMENTARY MATERIAL
Modolo et al. Assessing the potential of tropical secondary forests in the Central Amazon for timber production
Uses of commercial species included in the study based on the use experience of local communities.

Location of the secondary forest of FAEXP/UFAM (USF), the secondary forest Carapanã-Açu Farm (CSF) and the sampled area (plots) in each secondary forest.

Temporal analysis of forest cover in the secondary forest of FAEXP/UFAM - USF and in the secondary forest of Carapanã-Açu Farm - CSF.

Dead individuals of Croton matourensis (A) and Vismia cayennensis (B) observed in the forests.

Publication Dates
-
Publication in this collection
17 Mar 2025 -
Date of issue
2025
History
-
Received
04 Jan 2024 -
Accepted
12 Dec 2024